Тело гидры имеет вид продолговатого мешочка, стенки которого состоят из двух слоёв клеток — эктодермы и энтодермы .
Между ними лежит тонкая студенистая неклеточная прослойка — мезоглея , служащая опорой.

Эктодерма формирует покров тела животного и состоит из нескольких видов клеток: эпителиально-мускульные , промежуточные и стрекательные .
Самые многочисленные из них — эпителиально-мускульные.

Эктодерма

эпителиально-мускульная клетка
За счёт мускульных волоконец , лежащих в основании каждой клетки, тело гидры может сокращаться, удлиняться и изгибаться.
Между эпителиально-мускульными клетками находятся группы мелких, округлых, с большими ядрами и небольшим количеством цитоплазмы клеток, называемых промежуточными .

При повреждении тела гидры, они начинают усиленно расти и делиться. Они могут превращаться в остальные типы клеток тела гидры, кроме эпителиально-мускульных.
В эктодерме находятся стрекательные клетки , служащие для нападения и защиты. В основном они расположены на щупальцах гидры. Каждая стрекательная клетка содержит овальную капсулу, в которой свёрнута стрекательная нить.
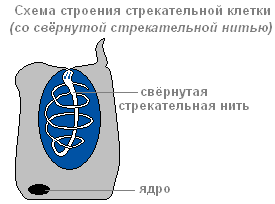
Строение стрекательной клетки со свернутой стрекательной нитью
Если добыча или враг прикоснётся к чувствительному волоску, который расположен снаружи стрекательной клетки, в ответ на раздражение стрекательная нить выбрасывается и вонзается в тело жертвы.
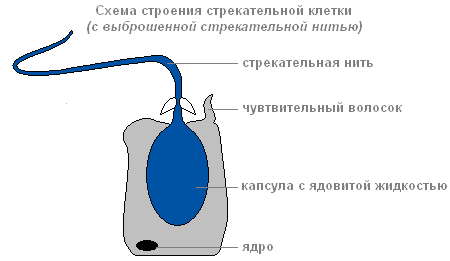
Строение стрекательной клетки с выброшенной стрекательной нитью
По каналу нити в организм жертвы попадает вещество, способное парализовать жертву.
Существует несколько типов стрекательных клеток. Нити одних пробивают кожные покровы животных и вводят в их тело яд. Нити других обвиваются вокруг добычи. Нити третьих — очень клейкие и прилипают к жертве. Обычно гидра «стреляет» несколькими стрекательными клетками. После выстрела стрекательная клетка погибает. Новые стрекательные клетки формируются из промежуточных .
Строение внутреннего слоя клеток
Энтодерма выстилает изнутри всю кишечную полость. В её состав входят пищеварительно-мускульные и железистые клетки.

Энтодерма
Пищеварительная система
Пищеварительно-мускульных клеток больше других. Мускульные волоконца их способны к сокращению. Когда они укорачиваются, тело гидры становится более тонким. Сложные движения (передвижение «кувырканием»), происходит за счёт сокращений мускульных волоконцев клеток эктодермы и энтодермы.
Каждая из пищеварительно-мускульных клеток энтодермы имеет 1-3 жгутика. Колеблющиеся жгутики создают ток воды, которым пищевые частички подгоняются к клеткам. Пищеварительно-мускульные клетки энтодермы способны образовывать ложноножки , захватывать и переваривать в пищеварительных вакуолях мелкие пищевые частицы.
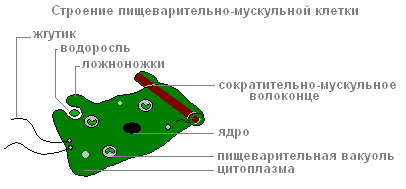
Строение пищеварительно-мускульной клетки
Имеющие в энтодерме железистые клетки выделяют внутрь кишечной полости пищеварительный сок, который разжижает и частично переваривает пищу.

Строение желистой клетки
Добыча захватывается щупальцами с помощью стрекательных клеток, яд которых быстро парализует мелких жертв. Координированными движениями щупалец добыча подносится ко рту, а затем с помощью сокращений тела гидра «надевается» на жертву. Пищеварение начинается в кишечной полости (полостное пищеварение ), заканчивается внутри пищеварительных вакуолей эпителиально-мускульных клеток энтодермы (внутриклеточное пищеварение ). Питательные вещества распределяются по всему телу гидры.
Когда в пищеварительной полости оказываются остатки жертвы, которые невозможно переварить, и отходы клеточного обмена, она сжимается и опорожняется.
Дыхание
Гидра дышит растворённым в воде кислородом. Органов дыхания у неё нет, и она поглощает кислород всей поверхностью тела.
Кровеносная система
Отсутствует.
Выделение
Выделение углекислого газа и других ненужных веществ, образующихся в процессе жизнедеятельности, осуществляется из клеток наружного слоя непосредственно в воду, а из клеток внутреннего слоя — в кишечную полость, затем наружу.
Нервная система
Под кожно-мускульными клетками располагаются клетки звездчатой формы. Это нервные клетки (1). Они соединяются между собой и образуют нервную сеть (2).
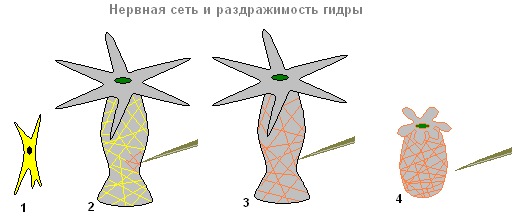
Нервная система и раздражимость гидры
Если дотронутся до гидры (2), то в нервных клетках возникает возбуждение (электрические импульсы), которое мгновенно распространяется по всей нервной сети (3) и вызывает сокращение кожно-мускульных клеток и всё тело гидры укорачивается (4). Ответная реакция организма гидры на такое раздражение — безусловный рефлекс .
Половые клетки
С приближением холодов осенью в эктодерме гидры из промежуточных клеток образуются половые клетки.
Различают два вида половых клеток: яйцевые, или женские половые клетки, и сперматозоиды, или мужские половые клетки.
Яйца находятся ближе к основанию гидры, сперматозоиды развиваются в бугорках, расположенных ближе к ротовому отверстию.
Яйцевая клетка гидры похожа на амёбу. Она снабжена ложноножками и быстро растет, поглощая соседние промежуточные клетки.

Строение яйцевой клетки гидры

Строение сперматозоида гидры
Сперматозоиды по внешнему виду напоминают жгутиковых простейших. Они покидают тело гидры и плавают с помощью длинного жгутика.
Оплодотворение. Размножение
Сперматозоид подплывает к гидре с яйцевой клеткой и проникает внутрь нее, причем ядра обеих половых клеток сливаются. После этого ложноножки втягиваются, клетка округляется, на ее поверхности выделяется толстая оболочка — образуется яйцо. Когда гидра погибает и разрушается, яйцо остается живым и падает на дно. С наступлением тёплой погоды живая клетка, находящаяся внутри защитной оболочки, начинает делиться, образующиеся клеточки располагаются в два слоя. Из них развивается маленькая гидра, которая выходит наружу через разрыв оболочки яйца. Таким образом, многоклеточное животное гидра в начале своей жизни состоит всего из одной клетки — яйца. Это говорит о том, что предки гидры были одноклеточными животными.
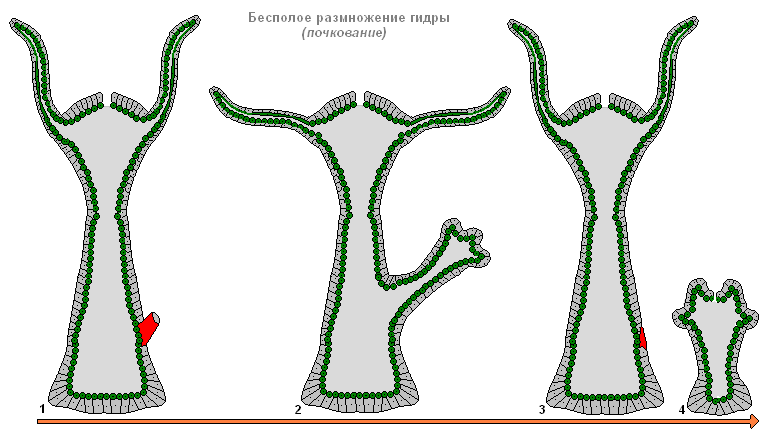
Бесполое размножение гидры
При благоприятных условиях гидра размножается бесполым путём. На теле животного (обычно в нижней трети туловища) образуется почка, она растет, затем формируются щупальца и прорывается рот. Молодая гидра отпочковывается от материнского организма (при этом материнский и дочерний полипы прикрепляются щупальцами к субстрату и тянут в разные стороны) и ведет самостоятельный образ жизни. Осенью гидра переходит к половому размножению. На теле, в эктодерме закладываются гонады - половые железы, а в них из промежуточных клеток развиваются половые клетки. При образовании гонад гидр формируется медузоидный узелок. Это позволяет предполагать, что гонады гидры - сильно упрощенные споросаки, последний этап в ряду преобразования утраченного медузоидного поколения в орган. Большинство видов гидр раздельнополы, реже встречается гермафродитизм. Яйцеклетки гидр быстро растут, фагоцитируя окружающие клетки. Зрелые яйцеклетки достигают диаметра 0,5-1 мм. Оплодотворение происходит в теле гидры: через специальное отверстие в гонаде сперматозоид проникает к яйцеклетке и сливается с ней. Зигота претерпевает полное равномерное дробление, в результате которого образуется целобластула. Затем в результате смешанной деламинации (сочетание иммиграции и деламинации) осуществляется гаструляция. Вокруг зародыша формируется плотная защитная оболочка (эмбриотека) с выростами-шипиками. На стадии гаструлы зародыши впадают в анабиоз. Взрослые гидры погибают, а зародыши опускаются на дно и зимуют. Весной продолжается развитие, в паренхиме энтодермы путем расхождения клеток образуется кишечная полость, затем формируются зачатки щупалец, и из-под оболочки выходит молодая гидра. Таким образом, в отличие от большинства морских гидроидных, у гидры отсутствуют свободноплавающие личинки, развитие у неё прямое.
Регенерация
Гидра обладает очень высокой способностью к регенерации. При разрезании поперек на несколько частей каждая часть восстанавливает «голову» и «ногу», сохраняя исходную полярность - рот и щупальца развиваются на той стороне, которая была ближе к оральному концу тела, а стебелек и подошва - на аборальной стороне фрагмента. Целый организм может восстанавливаться из отдельных небольших кусочков тела (менее 1/100 объёма), из кусочков щупалец, а также из взвеси клеток. При этом сам процесс регенерации не сопровождается усилением клеточных делений и представляет собой типичный пример морфаллаксиса.
Передвижение
В спокойном состоянии щупальца вытягиваются на несколько сантиметров. Животное медленно водит ими из стороны в сторону, подстерегая добычу. При необходимости гидра может медленно передвигаться.
«Шагающий» способ передвижения

«Шагающий» способ передвижения гидры
Изогнув своё тело (1) и прикрепившись щупальцами к поверхности предмета (субстрата), гидра подтягивает к переднему концу тела подошву (2). Затем шагающее движение гидры повторяется (3,4).
«Кувыркающий» способ передвижения
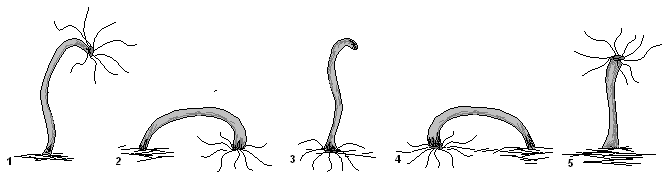
«Кувыркающий» способ передвижения гидры
В другом случае она словно через голову кувыркается, поочерёдно прикрепляясь к предметам то щупальцами, то подошвой (1-5).
4. Размножение и развитие
5. Рост и регенерация
6. Продолжительность жизни
7. Симбионты
8. История открытия и изучения
9. Гидра как модельный объект
Миграция и обновление клеток
В норме у взрослой гидры клетки всех трех клеточных линий интенсивно делятся в средней части тела и мигрируют к подошве, гипостому и кончикам щупалец. Там происходит гибель и слущивание клеток. Таким образом, все клетки тела гидры постоянно обновляются. При нормальном питании «избыток» делящихся клеток перемещается в почки, которые обычно образуются в нижней трети туловища.
Регенерационная способность
Гидра обладает очень высокой способностью к регенерации. При разрезании поперек на несколько частей каждая часть восстанавливает «голову» и «ногу», сохраняя исходную полярность рот и щупальца развиваются на той стороне, которая была ближе к оральному концу тела, а стебелек и подошва на аборальной стороне фрагмента. Целый организм может восстанавливаться из отдельных небольших кусочков тела, из кусочков щупалец, а также из взвеси клеток. При этом сам процесс регенерации не сопровождается усилением клеточных делений и представляет собой типичный пример морфаллаксиса.
Гидра может регенерировать из взвеси клеток, полученных путем мацерации. В экспериментах показано, что для восстановления головного конца достаточно образования агрегата из примерно 300 эпителиально-мускульных клеток. Показано, что регенерация нормального организма возможна из клеток одного слоя.
Опыты по изучению регенерации и модели регенерации
Уже ранние опыты Трамбле показали, что при регенерации сохраняется полярность фрагмента. Если разрезать тело гидры поперек на несколько цилиндрических фрагментов, то на каждом из них ближе к бывшему оральному концу регенерируют гипостом и щупальца, а ближе к бывшему аборальному полюсу подошва. При этом у тех фрагментов, которые располагались ближе к «голове», быстрее регенерирует «голова», а у располагавшихся ближе к «ноге» «нога».
Позднее опыты по изучению регенерации были усовершенствованы в результате применения методики сращивания фрагментов разных особей. Если вырезать из боковой стороны туловища гидры фрагмент и срастить его с телом другой гидры, то возможны три исхода опыта: 1) фрагмент полностью сливается с телом реципиента; 2) фрагмент образует выступ, на конце которого развивается «голова»; 3) фрагмент образует выступ, на конце которого образуется «нога». Выяснилось, что процент образования «голов» тем выше, чем ближе к «голове» донора взят фрагмент для пересадки и чем дальше от «головы» реципиента он помещен. Эти и аналогичные опыты привели к постулированию существования четырех веществ-морфогенов, регулирующих регенерацию активатора и ингибитора «головы» и активатора и ингибитора «ноги». Эти вещества, согласно данной модели регенерации, образуют концентрационные градиенты: в районе «головы» у нормального полипа максимальна концентрация как активатора, так и ингибитора головы, а в районе «ноги» максимальна концентрация и активатора, и ингибитора ноги.
Эти вещества действительно были обнаружены. Активатор головы пептид из 11 аминокислот, активный в пикомолярной концентрации. У человека он присутствует в гипоталамусе и кишечнике и в той же концентрации обладает нейротрофичесским действием. У гидры и млекопитающих этот пептид обладает также митогенным действием и влияет на клеточную дифференцировку.
Активатор ноги тоже пептид с молекулярной массой, близкой к 1000 Да. Ингибиторы головы и ноги низкомолекулярные гидрофильные вещества небелковой природы. В норме все четыре вещества выделяются нервными клетками гидры. Активатор головы имеет большее время полужизни, чем ингибитор и медленнее диффундирует, так как связан с белком-носителем. Ингибитор головы в очень низкой концентрации подавляет выделение активатора, а в 20 раз большей концентрации своё собственное выделение. Ингибитор ноги также ингибирует выделение активатора ноги.
Молекулярные механизмы регенерации
Получение «безнервных» гидр
При регенерации, как и при росте и бесполом размножении, эпителиально-мускульные клетки делятся самостоятельно, причем клетки эктодермы и энтодермы две независимые клеточные линии. Остальные типы клеток развиваются из промежуточных. Убив делящиеся промежуточные клетки высокой дозой радиации или колхицином, можно получить «безнервных», или эпителиальных гидр они продолжают расти и почковаться, но отделяющиеся почки лишены нервных и стрекательных клеток. Культуру таких гидр удается поддерживать в лаборатории с помощью «насильственного» кормления.
Первым смог разглядеть и описать гидру натуралист А. Левенгук, который изобрел микроскоп. Этот ученый был самым значительным натуралистом XVII-XVIII веков.
Рассматривая водные растения при помощи своего примитивного микроскопа, Левенгук заметил странное существо, у которого имелись руки «в виде рогов». Ученый даже пронаблюдал за почкованием этих существ и разглядел их стрекательные клетки.
Строение пресноводной гидры
Гидра относится к кишечнополостным животным. Ее тело имеет трубковидную форму, в передней части находится ротовое отверстие, которое окружено венчиком, состоящим из 5-12 щупалец.
Под щупальцами тело гидры сужается и получается шейка, которая отделяет туловище от головы. Задняя часть тела сужена в стебелек или ножку, с подошвой на конце. Когда гидра сытая, ее тело в длине не превышает 8 миллиметров, а если гидра голодная, тело значительно длиннее.
Как у всех представителей кишечнополостных, тело гидры образовано двумя слоями клеток.
Внешний слой состоит из разнообразных клеток: одни клетки используются для поражения добычи, другие клетки обладают сократимостью, а третьи выделяют слизь. А во внешнем слое имеются нервные клетки, которые формируют сеть, покрывающую тело гиды.
Гидра является одним из немногих представителей кишечнополостных, который обитает в пресной воде, а большая часть этих существ живут в морях. Средой обитания гидр являются разнообразные водоемы: озера, пруды, канавы, речные заводи. Они селятся на водных растениях и корешках ряски, которая покрывает ковром все дно водоема. Если вода чистая и прозрачная, то гидры селятся на камнях у самого берега, образуя порой бархатный ковер. Гидры любят свет, поэтому отдают предпочтение неглубоким местам рядом с берегами. Эти существа могут различать направление света и двигаться к его источнику. Если гидры живут в аквариуме, они всегда перебираются в его освещенную часть.

Если в сосуд с водой поместить водные растения, то можно заметить, как по их листьям и стенкам сосуда ползают гидры. На подошве гидры имеется клейкое вещество, которое помогает ей прочно крепиться к водным растениям, камням и стенкам аквариума, оторвать гидру с ее места довольно сложно. Изредка гидра передвигается в поиске пище, это можно наблюдать в аквариумах, когда на стеке остается след в том месте, где гидра сидела. За несколько дней эти существа перемещаются не более чем на 2-3 сантиметра. Во время движения, гидра крепится к стеклу щупальцем, отрывает подошву и перетаскивает ее на новое место. Когда подошва крепится к поверхности, гидра выравнивается и снова опирается на свой щупалец, совершая шаг вперед.
Этот способ перемещения схож с передвижением гусениц бабочек пядениц, которых часто называют «землемерами». Но гусеница подтягивает заднюю часть к передней, а затем снова перемещает переднюю часть. А гидра во время передвижения каждый раз переворачивается через голову. Так гидра передвигается достаточно быстро, но имеется и другой, более медленный способ перемещения – когда гидра скользит на своей подошве. Некоторые особи могут открепляться от субстрата и плавать в воде. Они расправляют свои щупальца и мелено опускаются на дно. А вверх гидры поднимаются при помощи пузырька газа, формирующегося на подошве.

Как питаются пресноводные гидры
Гидры являются хищными существами, они питаются инфузориями, циклопами, мелкими рачками – дафниями и другой мелкой живностью. Иногда они поедают более крупных жертв, например небольших червей или личинок комара. Гидры даже могут наносить ущерб рыбоводческим прудам, поскольку они поедают только что проклюнувшихся из икры рыб.
Как охотится гидра, можно легко проследить в аквариуме. Она широко расставляет щупальца, которые формируют сеть, при этом она висит щупальцами вниз. Если наблюдать за гидрой, можно заметить, что ее тело, медленно раскачиваясь, описывает передней частью круг. Проплывающая мимо жертва задевается о щупальца, пытается освободиться, но затихает, поскольку стрекательные клетки парализуют ее. Гидра подтягивает добычу ко рту и начинает поедать.
Если охота успешная, гидра раздувается от количества съеденных рачков, а их глазки просвечиваются сквозь ее тело. Гидра может съесть добычу, превышающую ее по размеру. Рот гидры способен широко раскрываться, а тело значительно растягиваться. Порой изо рта гидры торчит часть жертвы, которая не поместилась вовнутрь.

Размножение гидры пресноводной
Если питания достаточно, гидры быстро размножаются. Размножение происходит почкованием. Процесс роста почки от крохотного бугорка до сформировавшейся особи занимает несколько дней. Нередко на теле гидры образуются несколько почек, в то время, пока молодая особь не отделилась от материнской гидры. Таким образом, у гидр происходит бесполое размножение.
Осенью, когда температура воды понижается, гидры могут размножаться и половым способом. На теле гидры образуются половые железы в виде вздутий. В одних вздутиях формируются мужские половые клетки, а в других — яйцевые клетки. Мужские половые клетки свободно плавают в воде и проникают в полость тела гидр, оплодотворяя неподвижные яйцеклетки. Когда образуются яйца, гидра, как правило, погибает. При благоприятных условиях из яиц выходят молодые особи.
Регенерация у пресноводной гидры
У гидр проявляется поразительная способность к регенерации. Если гидру разрезать пополам, то в нижней части быстро отрастут новые щупальца, а на верхней части – подошва.
В XVII веке голландский ученый Трамбле проводил с гидрами интересные эксперименты, в результат которых ему не просто удавалось выращивать новых гидр из кусочков, но и сращивать различные половинки гидр, получать семиглавых полипов и выворачивать их тела наизнанку. Когда был получен семиглавый полип, похожий на гидру из Древней Греции, этих полипов и стали называть гидрами.
Как вы уже поняли из предшествующего изложения, к регенерации способны многие представители животного царства. Но форма и масштаб восстановительного роста у разных животных могут серьезно различаться. В этой главе нам предстоит познакомиться с четырьмя известными биологами, которым мы в значительной мере обязаны нашими знаниями о восстановлении утраченных органов. Каждый из этих ученых выбрал свой особый путь изучения проблемы регенерации, и вам станет понятно, что какого-то одного пути решения проблемы не существует. Понимание механизма регенерации может складываться лишь из тщательного сопоставления информации, получаемой с применением самых разных экспериментальных подходов.
ЭЛЛИСОН БЕРНЕТТ. РЕГЕНЕРАЦИЯ У ГИДР
Эллисон Бернетт преподает в Северо-западном университете в Эванстоне, штат Иллинойс. Большую часть своей научной деятельности он посвятил исследованию клеточной организации и процессов роста v гидр (Hydra ), относящихся к той же группе беспозвоночных животных, что и медузы, актинии и кораллы. Как еще в 1740 году впервые отметил Трамбле, регенерационная способность гидр не уступает по интенсивности регенерационной способности планарий. Поэтому не удивительно, что именно исследования регенерации у гидр и планарий явились предметом сотен научных сообщений на многих языках мира. Гидры - самый распространенный объект для постановки опытов как с учебной, так и с исследовательской целями.
Эти внешне напоминающие растения животные обычно обитают в прудах, прикрепившись к какому-нибудь водному растению или камню при помощи клеточного диска (подошвы), находящегося у основания тела. На противоположном («головном») конце трубкообразного тела гидры расположен рот, открывающийся в мешковидную пищеварительную полость. Он окружен венчиком щупалец (от шести до десяти), непрерывно двигающихся в поисках пищи. Одним из способов размножения гидр является почкование - образование небольших выпячиваний, или почек, в нижней части тела беспозвоночного. Постепенно на растущих почках формируются щупальца и другие типичные для гидр органы. Затем дочерняя особь отделяется от родительской и начинает самостоятельную жизнь. Образующиеся почки придают гидре обличье многоголового существа. Высокая регенерационная способность животного послужила основанием для того, чтобы назвать его по имени древнегреческого мифологического чудовища девятиголовой Гидры, способной легко восстанавливать головы, отсеченные в схватке (рис. 33). У гидры и родственных ей животных стенка тела состоит из клеток, покрывающих тело снаружи (эктодермы) и обладающих способностью к сокращению, и клеток, выстилающих пищеварительную полость (энтодермы); пространство между этими двумя слоями заполняет тонкий слой студенистого вещества, называемого мезоглеей. Наиболее часто встречающиеся гидры в длину не превышают 30 миллиметров.
Э. Бернетт изучал различные свойства гидр. Первые свои работы он посвятил исследованию структуры и функции специализированных клеток этих необычных животных: нервных клеток, которые создают сеть нервных образований в стенке тела, характерную для всех кишечнополостных; железистых клеток, входящих в состав энтодермы и секретирующих пищеварительные ферменты; а также расположенных на щупальцах стрекательных клеток, способных выбрасывать скрученную нить с ядом, парализующим мелких животных, добычу гидры, и в целях защиты. Кроме перечисленных во многих местах тела гидры можно обнаружить небольшие клетки, лишенные каких-либо специальных функций; их обозначают как промежуточные, или «интерстициальные», клетки («И-клетки»),
Регенерация у гидры
Следующие свои эксперименты Бернетт посвятил изучению особых ростовых факторов, которые, как он считает, выделяются гидрой. В пользу существования таких веществ говорили наблюдения над характером регенерации у гидры, Бернетту и другим исследователям удалось обнаружить, что зона роста животного находится в стенке тела непосредственно под щупальцами. Постоянное образование новых клеток в этой зоне приводит к тому, что лежащие рядом зрелые клетки постепенно оттесняются в двух противоположных направлениях - в сторону щупалец и в сторону основания тела,- а новые клетки, дифференцируясь, замещают их. Когда «старые» клетки достигают концов тела гидры, они слущиваются в окружающую водную среду. Согласно изложенной гипотезе, в случае, если какие-либо из специализированных клеток погибают до окончания процесса миграции, их место занимают близлежащие И-клетки, которые подвергаются соответствующим изменениям и берут на себя функции замещаемых клеток. Последний способ клеточного замещения наблюдается весьма часто: стрекательные клетки постоянно расходуются в процессе захвата добычи, а железистые - в процессе пищеварения. В результате организм гидры подвергается практически непрерывному обновлению (и тем и другим путем), за что это животное не без оснований получило название «бессмертного».
В дополнение к регенерационным механизмам, действующим непрерывно, гидры восстанавливаются и будучи повреждены в результате экспериментальных воздействий. Эти животные способны не только регенерировать любую из утраченных частей, но и полностью восстанавливать тело из любого мельчайшего фрагмента, за исключением щупалец и подошвы. В процессе регенерационного роста отмечается явная полярность: при горизонтальном рассечении гидры пополам «головная» часть, имеющая щупальца, восстанавливает от раневой поверхности стебелек с подошвой, и наоборот. На первый взгляд, у гидры отмечается градиент свойств по линии щупальца - подошва, подобный тому, который был описан на примере планарий. Бернетт, однако, высказал иное предположение. Несколько ранее он вместе с другими исследователями пришел к заключению, что зона роста под щупальцами выделяет особое ростовое вещество, стимулирующее процесс клеточного деления. Теперь же Бернетт предположил, что в той же самой зоне вырабатывается и подавляющее рост вещество и что именно от сочетания этих двух факторов зависит процесс как нормального, так и регенерационного роста гидры.
Модель роста гидры
Для иллюстрации справедливости своих гипотез исследователи нередко прибегают к созданию моделей тех или иных процессов. Предложенная Бернеттом модель регуляции роста гидры (рис. 34) предполагает, что и стимулирующее, и подавляющее рост вещества медленно перемещаются от места своей выработки в сторону основания тела животного, причем подавляющее рост вещество состоит из «текучих» молекул, которые постепенно выходят из тела в окружающую среду.

Какие же особенности регенерации гидры дали Бернетту основание для формулировки принципов работы его модели? Прежде всего, характер регенерации после рассечения. В верхней части, несущей щупальца, продуцируется как подавляющее, так и стимулирующее рост вещества. Логично предположить, что тут будет происходить нейтрализация одного фактора другим. И действительно, мы не наблюдаем роста щупалец на рассеченном конце, наоборот, здесь начинает формироваться стебелек с подошвой и восстанавливается характерная для тела животного полярность. Рост «головы» на раневой поверхности нижней половины гидры подтверждает два других постулата гипотезы: во-первых, в этой половине гидры нет клеток, способных вырабатывать подавляющее рост вещество, и, во-вторых, большая его часть, которая должна была достигнуть этого отдела тела, уже вышла в окружающую среду.
Помимо полярности регенерационного роста гидры модель Бернетта объясняет и некоторые аспекты нормальной формы ее роста, в частности размножение путем почкования. С точки зрения гипотезы о наличии в теле гидры градиента жизненных процессов по линии «щупальца - подошва» трудно понять механизм почкования. Согласно градиентной модели регенерации у планарий, темп любых биологических процессов значительно выше в головном конце животного, а у гидры быстрый рост, необходимый для почкования, происходит на участке тела, весьма отдаленном от «головы». Но с другой стороны, именно теория Бернетта легко объясняет наблюдаемое в природе явление. Важно только помнить, что предполагаемый фактор подавления роста наделен повышенной «текучестью». Это создает в нижних отделах тела гидры избыток стимулирующего рост вещества, который и обеспечивает активное отрастание дочерних особей Б районе стебелька. В «почках» вскоре начинается самостоятельная выработка подавляющего рост вещества, чем и объясняется полярность тела вновь образующихся гидр.
Каковы наиболее важные особенности модели регуляции роста гидры, предложенной Бернеттом? Она объясняет, во-первых, как нормальную, так и восстановительную формы роста этих беспозвоночных при помощи одной универсальной теории, и, во-вторых, наблюдаемую полярность роста взаимодействием двух специфических химических факторов. Это чрезвычайно ценные идеи, но тем не менее модель Бернетта не дает окончательного ответа на все вопросы, связанные с регенерацией у гидры. Ее значение состоит прежде всего в том, что она может послужить основанием для проведения дальнейших экспериментальных исследований, которые в настоящее время проводятся как самим Бернеттом, так и другими учеными, интересующимися данной проблемой.
МАРКУС СИНГЕР. НЕРВЫ И РЕГЕНЕРАЦИЯ
О значении нервов на определенных этапах регенерации конечности у амфибий мы уже говорили. Маркус Сингер из Медицинской школы при Кливлендском университете, штат Огайо, был первым, кто заинтересовался взаимоотношением нервной ткани и процессов регенерации в самых различных аспектах этой проблемы.
В опытах по денервации конечностей тритона Сингер установил, что регенерация зависит от сохранности нерва вплоть до стадии образования хорошо сформированной бластемы культи. Серия дальнейших весьма интересных исследований позволила Сингеру вскрыть возможный путь влияния нервной ткани на восстановительный процесс. Он пришел к заключению, что нервная ткань выделяет какое-то активное вещество, необходимое для протекания регенерации. Сингер говорит о необходимости исследования этого «нейротропного» агента на молекулярном уровне.
Качество необходимой нервной ткани
Каждый нерв, находящийся в конечностях позвоночных животных, состоит как бы из двух частей. Одна из них - сенсорная (чувствительная) - переносит нервные импульсы от конечности к центральной нервной системе независимо от того, каков характер раздражения конечности. Вторая часть - двигательная, она переносит сигналы от центральной нервной системы к мускулатуре конечности, обеспечивая реакцию на различные виды раздражений. На первых порах Сингер пытался установить, участвуют ли обе части нерва в восстановлении конечности тритона. Для этого непосредственно перед ампутацией передней конечности тритона ученый рассекал либо все чувствительные окончания трех основных нервов конечности, либо все двигательные (рис. 35). Оказалось, что регенерация успешно протекает при обоих вариантах постановки опыта, то есть при сохранении либо двигательной, либо чувствительной иннервации. Отсюда можно сделать вывод, что влияние нервной ткани на регенерацию не является качественным, поскольку тип нервного волокна, остающегося в рассеченной конечности, никак не сказывается на ее регенерационной способности. Но что можно сказать о количественной стороне дела? Как сказывается на процессе регенерации количество сохраненной нервной ткани?

Количество необходимой нервной ткани
Анализ результатов предыдущих опытов показывает что для нормального восстановления конечности нет необходимости сохранять нервную ткань в обычном количестве. Ведь полное восстановление конечности без чувствительных либо двигательных нервных окончаний идет при явной утрате значительной частя нервов. Но, поскольку полностью денервированная конечность не способна к регенерации, существует, по-видимому, некое минимальное количество нервной ткани, необходимое для ее восстановительного роста. Сингер предложил схему экспериментов, с помощью которых можно было установить значение такого минимума.
Как чувствительные, так и двигательные окончания трех основных нервов конечности состоят из пучков с определенным количеством нервных волокон, связанных между собой соединительной тканью. На первом этапе опыта устанавливали количество волокон в каждой из частей этих трех нервов. Подготовленные для микроскопирования препараты поперечных срезов нетронутых нервов окрашивали таким образом, чтобы можно было подсчитать число волокон как в чувствительном, так и в двигательном компонентах. При различных вариантах рассечения нервов у экспериментальных животных легко определить число оставшихся нервных элементов - для этого надо только вычесть число рассеченных из уже известного числа волокон данного нерва. Результаты были довольно интересные. Если в конечности оставалось более 1298 нервных волокон, регенерация протекала нормально, если их количество становилось ниже 793, регенерация не происходила. Если же число сохраненных нервных волокон лежало в пределах от 793 до 1298, восстановление конечности иногда происходило, а иногда нет. Таким образом, регенерацию обеспечивает некое среднее число нервных волокон (793-1298), так называемый пороговый уровень.
Логично предположить, что отсутствие способности к регенерации конечностей у того или иного животного может быть связано с недостижением порогового числа нервных волокон. Но дальнейшие эксперименты Сингера показали, что способность к регенерации определяется все же не общим числом оставшихся после ампутации нервных волокон. К этому заключению он пришел, сравнив число нервных волокон в конечностях животных ряда видов. У неспособных к регенерации животных, таких, как мыши или взрослые лягушки, полученные цифры были значительно ниже порогового числа у тритонов. Но подсчет числа нервных волокон у Xenopus , южноафриканской шпорцевой лягушки, неожиданно показал, что столь же низкое число нервных волокон у этих животных сочетается с хорошо выраженной регенерационной способностью, которая проявляется и во взрослом состоянии (рис. 36).

Это противоречие было разрешено, когда дополнительно к подсчету волокон определили размер нервов конечностей у представителей различных групп животных. При этом оказалось, что нервные волокна у Xenopus значительно превышают по диаметру такие же волокна у мышей и взрослых лягушек других видов. Вследствие этого степень иннервации конечности Xenopus значительно выше, чем у сравниваемых видов животных со сниженной регенерационной способностью. Под влиянием полученных данных пришлось несколько изменить концепцию порогового уровня. Теперь она гласит, что способностью к регенерации обладают конечности животных, у которых обеспечивается определенный уровень снабжения ампутированной зоны общим количеством нервной ткани, или нейроплазмы .
По мнению Сингера, концепция порогового уровня удачно объясняет, почему в процессе эволюции снижается способность к восстановлению конечностей, несмотря на явную эволюционную «полезность» регенерации. Он утверждает, что по мере усложнения центральной нервной системы происходило постепенное снижение количества нервной ткани в конечностях. В связи с этим у высших позвоночных постулируемый его теорией пороговый уровень иннервации конечностей не достигается. При этом Сингер считает, что природа не зря поступилась способностью к восстановлению конечностей, пойдя по пути все большего усовершенствования центральной нервной системы. Приобретение способности принимать быстрые решения, позволяющие животному эффективно защищаться от врагов, обладает в эволюционном плане большей ценностью, чем способность к отрастанию утраченных частей тела.
Каким образом нервы стимулируют рост тканей?
Следующим этапом было изучение механизмов влияния нервной ткани на регенерацию. Сингер предположил, что на ранних стадиях регенерации регулирующим воздействием обладает определенное химическое вещество, выделяемое нервами. Денервация конечности саламандр передили во время образования бластемы останавливает регенерацию из-за того, что прекращается выработка этого вещества. А что, если после денервации каким-то образом компенсировать отсутствие предполагаемого химического фактора-регулятора? Самым трудным при проведении этих опытовоказалось найти способ обработки денервированных регенерирующих конечностей саламандр различными химическими веществами. Пробовали наносить реагенты непосредственно на поверхность культи или вводить их в ткань с помощью шприца. Но ни в том, ни в другом случае возобновления регенерации денервированной культи не происходило. Нельзя было исключить и той возможности, что содержащийся в используемых препаратах химический стимулятор просто не достигал бластемы, как это бывает при его выделении нервами в естественных условиях. Для решения этой задачи Сингер предложил специальный аппарат, который должен был самым близким образом дублировать нормальную деятельность нервов, постепенно выделяя испытываемые вещества непосредственно в бластему конечности. Подобный процесс называется инфузией, в связи с чем изобретение было названо микроинфузионным аппаратом Сингера.
Микроинфузия
Предложенный Сингером аппарат предназначен для непрерывного протекания небольших объемов жидкости через конечности тритонов, денервированные на ранней стадии регенерации. Действие аппарата основано на вращении часового механизма, которое преобразуется в поступательное движение винта. Винт, в свою очередь, приводит в движение поршень небольшого подкожного шприца, подающего раствор в тонкую пластиковую трубочку, вставленную на место иглы. Свободный конец трубочки заканчивается стеклянным капилляром, который вводится в плечевую область тритона после обезболивания животного и затем проникает в ткани культи и бластемы. Винтовая часть механизма может быть соединена с подвижной пластинкой, нажимающей из поршни ряда шприцов,- такая модификация дает возможность проводить одновременнуюинфузию нескольким тритонам (рис. 37).

Тритонов держат под наркозом до пяти часов и за это время проводят инфузию различных химических веществ в регенерирующую конечность. Для того чтобы инфузия соответствовала естественному процессу высвобождения химических веществ тканью нерва, в конечность поступают минимальные объемы испытываемых растворов - около 0,001 миллилитра в час.
Сингер предположил, что наиболее вероятным веществом, оказывающим влияние на регенерационную способность, может быть медиатор (передатчик нервного напряжения) ацетилхолин. Для этого предположения имелось несколько причин. Во-первых, известно, что ацетилхолин высвобождается нервной тканью при передаче импульсов. Во-вторых, при определении содержания ацетилхолина в конечности на различных этапах регенерации было обнаружено, что на так называемых нервно-зависимых этапах его количество оказалось большим по сравнению с нормальной тканью. После формирования бластемы и во время фазы респециализации содержание ацетилхолина возвращалось к нормальному уровню (рис. 38).

В регенерирующие конечности тритонов на протяжении разных периодов времени вводили различные концентрации ацетилхолина. Казалось весьма вероятным, что инфузия ацетилхолина хотя бы в некоторых случаях обеспечит возможность регенерации денервированной конечности. Но ожидания не оправдались. Инфузия денервированных конечностей ни разу не привела к завершению процесса регенерации.
Несмотря на некоторое разочарование, вызванное результатом этих опытов, его все же следует считать ценным, поскольку он исключает один из возможных механизмов влияния нервной ткани на регенерацию и позволяет сосредоточить внимание исследователей на поисках альтернатив.
Молекулярная биология и регенерация конечностей
Природа вещества, с помощью которого нервная ткань оказывает влияние на регенерацию конечностей, до сих пор остается невыясненной. Исследователи, занимающиеся этим вопросом, недавно попытались расшифровать механизм воздействия нервов на регенерирующие клетки, надеясь таким образом определить участвующее в данном процессе вещество.
Когда конечность денервируется в стадии образования бластемы, процесс регенерации прерывается, указывая тем самым на остановку жизнедеятельности клеток бластемы. Вполне логично предположить, что денервация воздействует на одну из наиболее важных функций клеток, а именно на процесс синтеза белков.
Детали процесса синтеза белков в клетках описаны в любом руководстве по биологии, но вкратце их можно сформулировать следующим образом. Молекулы ДНК, располагающиеся в ядре каждой клетки, содержат закодированную информацию для синтеза различных белков. Они служат своего рода матрицей для формирования молекул информационной РНК, которые переносят указанную информацию к расположенным в цитоплазме клеток рибосомам. Именно здесь происходит процесс сборки белков из отдельных «строительных блоков», которыми являются аминокислоты. В опытах с использованием радиоактивных изотопов Сингер и его сотрудники пытались определить, какое влияние оказывает денервация конечности тритона в ранней фазе образования бластемы на синтез белка в клетках конечности. Они исходили из того, что при денервации данный тип клеточной активности должен останавливаться или хотя бы снижаться.
Аминокислоты, подобно многим другим химическим веществам, могут быть «помечены», если некоторые из элементов заменить на радиоактивные. По интенсивности включения меченых аминокислот в молекулы белка можно определить уровень синтеза белка в клетках бластемы. Соответственно эффект денервации должен проявиться в изменении уровня включения меченых аминокислот в белки, синтезируемые клетками бластемы.
Итак, в ткань регенерирующих конечностей тритонов вводили меченые аминокислоты. Для выявления метки в данном случае применяли не авторадиографию, а другой метод. В разные сроки после введения меченых аминокислот получали бластему, растирали ее и выделяли белки. Пробы полученных материалов помещали в сцинтилляционный счетчик - прибор, позволяющий определить уровень радиоактивности, высвобождаемой определенным количеством белка за минуту.
Результаты проведенных опытов подтвердили исходную гипотезу: препарат белков клеток бластемы денервированных конечностей был значительно менее радиоактивным по сравнению с аналогичным препаратом из конечностей, где иннервация была сохранена. Таким образом, синтез белка в клетках бластемы действительно зависит от присутствия вещества, выделяемого нервной тканью. Для подтверждения этого вывода Сингер провел опыты по культивированию нервной ткани и материал из культур инфузировал в бластемы регенерирующих конечностей, которые были перед тем денервированы. Потом он повторил опыт с радиоизотопным исследованием синтеза белка клетками бластемы. После инфузии материала из культур нервной ткани клетки бластем денервированных конечностей включали меченые аминокислоты примерно с той же интенсивностью, что и клетки бластем с сохраненной иннервацией.
Так впервые был выявлен на молекулярном уровне эффект выделяемого нервной тканью вещества, влияющего на процесс регенерации конечности. Открытие, важное само по себе, но имеющее и более общее значение,- оно стимулировало интерес ученых к этому аспекту проблемы регенерации и заложило основу изучения специфических регуляторных механизмов данного биологического явления.
ЭЛИЗАБЕТ ХЕЙ. ИССЛЕДОВАНИЕ ПРОЦЕССОВ РЕГЕНЕРАЦИИ С ПОМОЩЬЮ ЭЛЕКТРОННОГО МИКРОСКОПА
Электронно-микроскопические исследования, проведенные Элизабет Хей в Гарвардской медицинской школе, внесли много нового в наше понимание судьбы отдельных клеток при регенерации. С помощью оптического микроскопа, как мы уже знаем, была определена общая последовательность изменений на уровне клеток, которые происходят в регенерирующих конечностях саламандр на стадиях дедифференцировки, образования бластемы и редифференцировки. Однако ограничение увеличений, достигаемых в оптическом микроскопе, оставляло многие вопросы без ответа. Так, например, при изучении регенерации у планарий микроскопия оказалась неспособной дать ответ, происходит ли формирование бластемы за счет миграции резервных клеток-необластов или за счет дедифференцировки зрелых клеток плоского червя. Тщательные электронно-микроскопические исследования, проведенные Э. Хей, позволили разрешить ряд важных проблем регенерации у беспозвоночных и позвоночных животных и получить много дополнительной информации.
Клетки регенерирующей конечности
При использовании обычного оптического микроскопа оставались неразрешенными три основных вопроса в изучении клеток регенерирующих конечностей саламандр. Первый из них относился к отдельным компонентам клетки, органоидам. Предполагалось, что органоиды клеток культи принимают активное участие в формировании бластемы. Но какие изменения претерпевают органоиды при дедифференцировке зрелых клеток? Детали этого процесса не поддаются изучению в оптическом микроскопе.
Второй вопрос вытекал из данных оптической микроскопии, показавшей, что клетки, составляющие бластему конечности, не несут никаких следов происхождения от тех или иных «родительских» клеток и по строению неотличимы друг от друга. Действительно ли клетки бластемы идентичны? На это указывали многие данные, но все же нельзя было исключить каких-то различий, невидимых в оптическом микроскопе. Требовались дополнительные исследования для того, чтобы доказать, что клетки бластемы действительно утратили все структурные признаки дифференцированных клеток.
Третья проблема касалась неясности в вопросе о том, какие именно ткани конечности подвергаются дедифференцировке и входят в состав бластемы. В особенности это относилось к утере специализации мышечной тканью культи. Микрофотографии, полученные в оптическом микроскопе, показывали, что рассеченные концы мускулатуры конечности становятся «растрепанными» в следующей за ампутацией стадии «демонтирования» и некоторые мышечные клетки в этом участке отделяются от основной мышечной массы, дедифференцируются и мигрируют к раневой поверхности.Тем не менее ряд исследователей придерживались точки зрения, что мышечная ткань не подвергается процессу дедифференцировки. Они считали, что после освобождения поврежденных концов нетронутой мускулатуры от клеточного детрита идет прямое отрастание новой мышечной ткани и проникновение мышц во вновь формируемую часть конечности. Проведенные Хей электронно-микроскопические наблюдения позволили более детально изучить строение цитоплазмы регенерирующих клеток и дали ответ на эти вопросы. Как вы уже, наверное, поняли, для этой цели использовался просвечивающий электронный микроскоп. Исследовали ультратонкие срезы нормальных и регенерирующих конечностей аксолотлей со специальным вниманием к структуре клеток мышц и хряща, поскольку эти клетки легко выявляются в зрелом состоянии по выделяемым ими специфическим веществам.
Прежде всего был установлен характер клеток двух упомянутых типов в неампутированной конечности. В цитоплазме зрелых хрящевых клеток при больших увеличениях хорошо просматривались многочисленные мембраны и рибосомы - мелкие внутриклеточные частицы, служащие для сборки белков из аминокислот. Рибосомы находились в тесной связи с мембранными структурами. Можете ли вы вспомнить другой случай выявления подобной картины? Да, нечто подобное мы уже видели на электронных микрофотографиях фибробластов, участвующих в процессе заживления ран. Матрикс, окружающий хрящевые клетки, содержит коллаген, как и ткани рубца, образуемые фибробластами, поэтому оба типа клеток синтезируют молекулы этого белка на рибосомах, связанных с мембранами. В хрящевых клетках нормальной конечности обнаруживается и комплекс Гольджи, который является типичным для железистых клеток. В зрелых мышечных клетках практически все пространство цитоплазмы занято пучками сократительного материала, поперечная исчерченность которого четко выявляется при увеличениях электронного микроскопа.
Электронные микрофотографии показали, что мышечные клетки подвергаются трансформации в тканях бластемы. На препаратах, полученных Хей в ранние стадии регенерации, в местах рассечения мышечной ткани среди множества ядер оставшейся неповрежденной мускулатуры были видны границы вновь образующихся клеток. Здесь же обнаруживались небольшие клетки, в каждой из них было по одному ядру. Позднее эти клетки, по-видимому, мигрировали к раневой поверхности конечности и становились клетками бластемы.
При исследовании в электронном микроскопе клетки ранней бластемы конечности аксолотля четко отличались от зрелых мышечных или хрящевых клеток (рис. 39). Например, цитоплазматические мембраны клеток бластемы были фрагментированными, а рибосомы свободно разбросаны по всей цитоплазме и не прикреплены к мембранам. Хотя комплекс Гольджи в клетках бластемы оставался различимым, он был гораздо меньше по сравнению с комплексом Гольджи в зрелых клетках хряща. Цитоплазма клеток бластемы была развита чрезвычайно слабо, зато ядра обладали гигантскими размерами и содержали четко выраженные ядрышки. Наконец, поскольку изучение ультраструктуры клеток бластемы не выявило даже следов хрящевого матрикса или мышечных фибрилл, было полностью подтверждено заключение, полученное при оптической микроскопии, об идентичности бластемных клеток.

На электронных микрофотографиях, сделанных в период редифференцировки, было видно, что «упростившиеся» органоиды клеток бластемы по мере восстановления конечности претерпевают постепенные изменения, характер которых определяется тем, какие именно специализированные клетки возникают на месте бластемы. В расположенных центрально клетках-предшественниках хряща постепенно «проявляются» цитоплазматические мембраны с прикрепившимися к ним рибосомами, комплекс Гольджи становится более выраженным, и вскоре вокруг клеток начинает обнаруживаться внеклеточный матрикс. В довольно поздних стадиях регенерации, когда уже четко видны границы восстанавливающихся костей, будущие мышечные клетки, расположенные в наружных частях бластемы, еще не имеют признаков респециализации. Но затем проступают и эти признаки, клетки удлиняются, а в цитоплазме начинает выявляться сократительный материал. Еще позже клетки сливаются и формируют типичную мышечную ткань (рис. 40). Таким образом, цитологическое исследование фазы респециализации регенерирующих конечностей аксолотля позволило ответить на все три вопроса, поставленных в начале раздела.

Электронная микроскопия и регенерация у планарий
Многие исследователи обнаруживали в оптическом микроскопе группы совершенно неспециализированных клеток, расположенные в разных участках тела плоских червей. Эти клетки не имели каких-либо четких отличий и различались только по характеру окраски их цитоплазмы некоторыми красителями. Поскольку они мигрировали в сторону раневых поверхностей и участвовали в формировании бластемы, их назвали резервными клетками (необластами). Считалось, что необласты распространены у всех видов плоских червей. Недавно Хей провела электронно-микроскопическое исследование этих резервных клеток у нормальных и регенерирующих планарий. Первое, что она обнаружила в клетках нормальных плоских червей,- это значительное число структурных деталей, указывающих на то, что исследуемые клетки не были в полном смысле слова неспециализированными. Большие увеличения электронного микроскопа позволили увидеть в этих клетках секреторные гранулы и структуры комплекса Гольджи - четкие «пограничные столбы» железистых клеток. Возникло предположение, что резервные клетки предназначены не столько для реакции на какие-то виды повреждений, сколько для определенной постоянной функции - выработки и выделения слизи. Слизь покрывает тело червя и позволяет ему с помощью мышечных сокращений перемещаться по самым разным поверхностям.
У регенерирующих планарий электронная микроскопия выявила своеобразные клеточные потоки, направляющиеся в сторону раневой поверхности. Однако в составе этих потоков были обнаружены не только железистые клетки, но и ряд других специализированных клеток. Позднее близ места рассечения червя мигрировавшие клетки постепенно утрачивали черты специализации, то есть дедифференцировались точно таким же образом, как клетки в регенерирующих конечностях земноводных. По достижении раневой поверхности все мигрирующие клетки оказывались полностью дедифференцированными и были готовы к формированию бластемы. Необласты, таким образом, были совершенно не нужны.
Как видите, электронная микроскопия во многих случаях подтверждает данные, основанные на наблюдениях в оптическом микроскопе. Таковы были результаты детального изучения клеток регенерирующей конечности. Но на примере с резервными клетками планарий электронно-микроскопические данные не совпали с результатами, полученными ранее с помощью менее совершенной техники. В связи с этим иногда приходится подвергать повторному исследованию объекты, казалось бы, давно изученные, после чего нередко происходит пересмотр установленных в науке положений.
РИЧАРД ГОСС. РАЗНООБРАЗИЕ РЕГЕНЕРАЦИОННЫХ СИСТЕМ
Ричард Госс работает в Браунском университете города Провиденс, штат Род-Айленд. Свою дгятельность он посвятил изучению проблемы регенерации различных органов у весьма широкого круга животных. Мы приведем здесь лишь две его работы, характеризующие чрезвычайную широту его исследовательских интересов. Речь пойдет о регенерации вкусовых усиков (мелких бакенбардоподобных чувствительных выростов, окружающих ротовое отверстие у представителей некоторых видов рыб) и крупных ветвящихся рогов у оленей и лосей, достигающих порой 130 сантиметров в длину.
Регенерация вкусовых усиков
Сом (по-английски «catfish», буквально «рыба-кошка») получил свое название именно благодаря чрезвычайно выраженным вкусовым усикам, очень напоминающим кошачьи усы. Д-р Госс обнаружил, что при отсечении у сома такого усика на его месте образуется бластема и утраченный отросток регенерирует. При микроскопическом исследовании структура усиков оказалась очень простой: каждый из них содержал нервы и кровеносные сосуды, основу органа составлял хрящевой стержень, а на верхушке под слоем эпидермиса находилась вкусовая почка.
Экспериментальное исследование этой миниатюрной регенерационной системы выявило ряд интересных фактов. Бластема, образующаяся после отсечения усика, формировалась только из дедифференцированных клеток хряща. Если через небольшой разрез у основания усика удаляли хрящевой стержень, а затем пересекали и сам усик, бластема не формировалась и отросток не регенерировал. Поскольку хрящевой стержень оказался необходимым для регенерации усика, логично было предположить, что если в один усик поместить несколько стержней (можно до четырех), то после ампутации усика с пересечением всех стержней появляющийся отросток будет содержать столько стержней, сколько их было в культе. Но эксперимент выявил в регенерирующем усике только один стержень. По-видимому, бластема вкусового усика «запрограммирована» на формирование нормального числа стержней в отростке, и присутствие дополнительных структур в культе не влияет на нормальный рост.
Регенерация рогов у оленей
В дальнейшем Госс сосредоточил свое внимание на изучении регенерации рогов у оленей. Периодическое естественное замещение этих структур являтся, по-видимому, единственным примером регенерации столь сложного органа у млекопитающих. Тем не менее он показывает, что и у теплокровных животных возможно восстановление крупных частей тела. Поэтому неудивителен большой интерес, проявляемый многими исследователями к изучению данной формы регенерации. В силу этого общий характер роста и судьба тех или иных клеток при восстановлении оленьих рогов, а также факт гормональной зависимости процесса регенерации изучены достаточно хорошо. Госсу, однако, удалось открыть в последнее время ряд новых способов воздействия природных сигналов на стимуляцию гормональной активности организма.
В ранний период роста у самцов оленя по обеим сторонам черепа, несколько выше и позади глаз, появляются небольшие костные выросты, или пеньки. Позднее на этих местах образуются мягкие округлые «роговые почки», которые затем удлиняются и ветвятся. Рост и развитие рога происходит из верхнего его конца, но окостенение хрящевых клеток осуществляется постепенно по мере удаления от основания рога снизу вверх. Подобный градиент дифференцировки тканей отмечается при каждом цикле регенерации рогов.
У самцов оленей, живущих в зоне умеренного пояса, сбрасывание и отрастание рогов происходит ежегодно, причем у разных видов наблюдаются довольно существенные вариации как в размере рогов, так и в интенсивности процесса их восстановления. Госс составил таблицы, которые показывают, что у более крупных животных рост рогов идет быстрее. У лосей, самых крупных представителей семейства оленей, рога могут достигать в длину 129,5 сантиметра и расти со скоростью 2,75 сантиметра в день (рис. 41, А, Б). У всех оленей по мере окостенения растущего рога пронизывающие его кровеносные сосуды перекрываются костной тканью, и кожа с короткими густыми волосами («вельвет»), покрывающая рога снаружи, лишаясь кровоснабжения, лопается и отпадает. Отпадение рогов, превратившихся в плотную компактную костную массу, происходит значительно позже, тогда, когда в месте соединения рогов и пеньков появляются клетки, разрушающие костные структуры. Ранки быстро заживают и рост рогов начинается заново. У большинства видов отпадение рогов происходит в конце зимы или весной, регенерацию можно видеть в летние месяцы, а сбрасывание «вельвета» проходит как раз перед периодом размножения, то есть осенью. Все эти процессы приводятся в движение зависящими от времени года колебаниями гормональной активности у оленей. Снижение количества гормона тестостерона весной стимулирует отпадение рогов и начало регенерации, а повышение его уровня осенью приводит к постепенному окостенению рогов и потере «вельвета».

Если вы живете в средней полосе, вы знаете, что в разные времена года длина светового дня различна. Цикл роста рогов и изменений гормонального уровня у оленей связан непосредственно с сезонными изменениями длины светового дня. Госс создал в своих экспериментах режим искусственного освещения, чтобы получить ответы на следующие вопросы; во-первых, можно ли изменить цикл смены рогов, удлиняя или укорачивая длительность искусственного освещения, и, во-вторых, будет ли происходить замещение рогов в условиях, когда длина светового дня будет постоянной на протяжении всего календарного года?
На первом этапе своих экспериментов он подвергал оленей воздействию «светового года» (полный годовой цикл возрастания и убывания длины светового дня), который на шесть месяцев расходился по фазе с обычным годом, то есть зимой дни становились длиннее, а летом - короче. Подопытных животных (пятнистых оленей - небольших, легкого сложения оленей, живущих в естественных условиях на Дальнем Востоке, но встречающихся в зоопарках всего мира) содержали в неотапливаемом помещении, где температура подвергалась естественным годовым изменениям. Специальный календарный механизм был соединен с осветительными приборами, с помощью которых поддерживалось «извращение времен года»: осенью по мере падения температуры длина светового дня постепенно увеличивалась, тогда как весной с потеплением световой день сокращался. При содержании в таких условиях регенерация рогов у пятнистого оленя приходилась на зимние месяцы, а сбрасывание - на осень. Животные подверглись полной адаптации к извращенным условиям освещения, во всяком случае, в том, что касалось роста и регенерации рогов.
Стремясь всесторонне проверить свою гипотезу, Госс в следующих опытах подвергал животных воздействию нескольких искусственных световых циклов на протяжении одного календарного года. Для этого календарный механизм настраивали так, чтобы он пропускал каждый второй день. Это обеспечило проведение за год двух годовых циклов изменения продолжительности светового дня. Если же механизм пропускал по два или три дня, годовые циклы повторялись трижды или четырежды в год. При помещении в такие условия пятнистые олени начинали терять рога дважды, трижды или четырежды в год в соответствии с числом экспериментальных годовых циклов. Из-за сокращения циклов длина отраставших рогов была, естественно, значительно меньше, чем при нормальном цикле (рис. 42, А, Б).

В следующем эксперименте Госс удлинил цикл светового года. Для этого календарный механизм дважды повторял каждый нормальный день, создав тем самым «двадцатичетырехмесячный год». Теперь ученый пытался получить ответ на следующие вопросы: продлится ли цикл роста рогов у содержащихся в таких условиях оленей все 24 месяца и скажется ли это на размерах рогов или нет? Ответ на первый вопрос оказался неодинаковым в опытах на оленях разного возраста: взрослые животные не изменяли цикла смены рогов, тогда как молодые олени легко приспосабливались к новой длительности «года», восстанавливая рога всего один раз за фактически два календарных года. Что же касается второго вопроса, то ни разу не наблюдалось отрастания рогов, превышающих по длине нормальные, хотя аномалии в характере их роста иногда были. Но во всех случаях искусственного изменения времен года наблюдаемые биологические эффекты не носили постоянного характера: большинство оленей по возвращении к естественному содержанию восстанавливали обычную цикличность роста рогов.
Может ли какой-нибудь искусственно созданный световой режим полностью приостановить смену рогов? С этой целью экспериментатор вообще исключал какие-либо колебания в продолжительности светового дня. На протяжении ряда лет специальную группу оленей содержали в условиях смены света и темноты ровно через 12 часов. Подобные условия были идентичны тем, которые наблюдаются на экваторе. Свет включали в 6 часов утра и гасили в 6 часов вечера. В этой группе большинство оленей вообще утратили способность к смене рогов, их регенерационный цикл был полностью нарушен. Кроме того, было установлено, что утеря цикла связана с постоянно повышенным уровнем тестостерона.
И наконец, в последнем варианте опытов Госс воздействовал на группу оленей с помощью постоянных на протяжении календарного года циклов, состоящих из неравных периодов света и темноты: восемь, шестнадцать или двадцать четыре часа света сопровождались соответственно шестнадцатью, восемью часами темноты или свет не выключали вообще. В каждом таком случае искусственно удлиненных или укороченных световых дней животные оказались способны с достаточно большой точностью определять фактический ход времени. Они сменяли рога один раз в год и удивительно близко к срокам, когда этот процесс протекает в естественных условиях. Эти результаты дают веские основания предполагать, что оленям свойствен внутренний ритм типа «биологических часов». (В предыдущих опытах Госса такой ритм подвергался адаптационным изменениям при искусственных извращениях годовых циклов или полностью нарушался при переводе животных в «экваториальные» условия освещения, когда свет и темнота чередовались каждые 12 часов.) Каков бы ни был физиологический механизм наблюдаемого внутреннего ритма, он зависит от главного фактора - неравенства продолжительности светлых и темных периодов в каждом 24-часовом цикле.
Из результатов, полученных другими исследователями, вытекает, что тип реакции, наблюдаемой у оленей, не исключение. У большого круга животных физиологические изменения и приспособительные реакции тесно связаны со сменой дня и ночи, сменой времени года, чередованием приливов и отливов. Во многих случаях, когда животных извлекали из естественной среды обитания и лишали многочисленных ее «сигналов», они тем не менее сохраняли чувство времени и соответственно поддерживали свои нормальные биологические циклы.
Результаты опытов Госса по регенерации рогов у оленей позволяют предположить, что сходным образом может осуществляться и регуляция других форм восстановительных процессов. Действительно, в последнее время наши знания о явлениях, относящихся к функционированию «биологических часов», непрерывно расширяются. И по-видимому, недалек тот день, когда мы узнаем, каким образом приводится в движение этот удивительный часовой механизм природы.
Биологи, с которыми мы познакомились в этой главе, продолжают свои исследования по регенерации. Эллисон Бернетт в экспериментах на гидрах уточнил механизмы регуляции роста у этих кишечнополостных животных. Кроме того, он пытается приложить некоторые из своих теорий к анализу регенерации у позвоночных. Поскольку основные закономерности жизнедеятельности клеток у животных различных эволюционных групп чрезвычайно близки, естественно, что специалисты по росту и развитию какой-то одной группы животных стремятся распространить свои выводы на другие. Маркус Сингер, установив, что высвобождаемый нервной тканью фактор может влиять на клеточные механизмы синтеза белка несколькими путями, занялся поисками биохимической «мишени» этого вещества в клетках бластемы конечности. Среди многих аспектов нейробиологии, которыми он интенсивно занимается, его особое внимание привлекает изучение структуры и функций миелиновой оболочки аксонов. Что касается Элизабет Хей, то ее мастерство электронного микроскописта служит теперь уже не только задачам изучения регенерации. Она исследовала тонкую структуру самых разных эмбриональных клеток - в частности, клеток сердца и хрусталика куриного эмбриона - и сопоставила свои наблюдения с функциями этих клеток в процессе эмбрионального развития. Ричард Госс в настоящее время наибольшее внимание уделяет таким процессам компенсаторной регенерации, как восстановление ткани печени и почки млекопитающих после удаления соответствующего органа. По мнению Госса, углубление знаний об этих процессах приведет к открытию специфических регуляторов роста тканей и органов у млекопитающих.
Работы этих ученых - и только что описанные, и осуществляемые ими в настоящее время - представляют, конечно, лишь часть той большой исследовательской деятельности, которая посвящена изучению регенерационных процессов. Только вместе взятые, они могут дать целостную картину каждого конкретного процесса. Но в общем перспектива представляется обнадеживающей. Регенерация стала важным разделом биологии развития - отрасли науки, занимающейся изучением закономерностей нормального и патологического роста, клеточной дифференцировки, экспериментальной эмбриологии и многими другими близкими проблемами. Благоприятные перспективы развития новой отрасли науки определяются еще и тем, что в последние годы в нее непрерывно вливаются новые энтузиасты изучения регенерации.
Движения гидры. Эпителиально-мускульные клетки эктодермы имеют во-локна, которые могут сокращаться. Если они сокращаются одновремен-но, укорачивается все тело гидры. Если сокращаются волокит в клетках с одной стороны, то в эту сторону гидра наклоняется. Благодаря работе этих волокон движутся щупальца гидры и перемещается все ее тело (рис. 13.4).
Реакции на раздражение гидры. Благодаря нервным клеткам, располо-женным в эктодерме, гидра воспринимает внешние раздражения: свет, прикосновения, некоторые химические вещества. Отростки этих клеток смыкаются между собой, образуя сетку. Так формируется самая простая по строению нервная система, называемая диффузной (рис. 13.5). Больше всего нервных клеток находится возле подошвы и на щупаль-цах. Проявлением работы нервной системы и эпителиально-мышечных клеток является безусловный рефлекс гидры — сгибание щупалец в от-вет на прикосновение.
| Рис. 13.4. Схема движении гидры |
 |
| Рис. 13.5. Нервная система гидры |
Во внешнем слое расположены также стрекательные клетки, содер-жащие капсулы со скрученной тоненькой трубочкой — стрекательной нитью. Наружу из клетки торчит чувствительный волосок. Достаточно его слегка коснуться, как нить выбрасывается из капсулы и вонзается в тело врага или добычи. К нему по стрекательной нити поступает яд, и животное погибает. Больше всего стрекательных клеток расположено в щупальцах.
Регенерация гидры. Маленькие округлые промежуточные клетки эктодер-мы способны превращаться в другие виды клеток. За счет их размноже-ния гидра быстро отстраивает поврежденную часть тела. Способность к регенерации этого животного поражает: когда гидру разделили на 200 частей, из каждой восстановилось целое животное!
Питание гидры. В эндодерме расположены железистые клетки и осна-щенные жгутиками пищеварительные клетки. Железистые клетки по-ставляют в кишечную полость вещества, называемые пищеваритель-ными соками. Эти вещества разрушают добычу, разлагая ее на микроскопические кусочки. С помощью жгутиков пищеварительные клетки подгоняют их к себе и захватывают, образуя псевдоподии. Внутреннюю полость гидры не случайно называют кишечной: в ней начинается переваривание пищи. Но окончательно пища расщепляет-ся в пищеварительных вакуолях пищеварительных клеток. Непе-реваренные остатки пищи удаляются наружу из кишечной полости через рот.
Выделение вредных веществ, образовавшихся в процессе жизнедея-тельности гидры, происходит через эктодерму в воду
Взаимодействие клеток. Среди клеток гидры лишь пищеваритель-ные переваривают пищу, но они обеспечивают питательными вещества-ми не только себя, но и все другие клетки. В свою очередь «соседи» создают наилучшие условия жизни для поставщиков питательных ве-ществ. Вспомните об охоте гидры — теперь вы сможете объяснить, как согласованная работа нервных, жалящих, эпителиально-мышечных и железистых клеток обеспечивает работой пищеварительные клетки. А эти клетки делятся с соседями результатами своего труда. Материал с сайта
Как размножается гидра? При бесполом размножении в результате деления промежуточных клеток образуется почка. Почка растет, на ней появляются щупальца, между ними прорывается рот. На противоположном конце формируется подошва. Маленькая гидра от-деляется от материнского организма, опускается на дно и начинает жить самостоятельно.
Размножается гидра и половым путем. Гидра — гермафродит: в од-них выступах ее эктодермы из промежуточных клеток формируются сперматозоиды, в других — яйцеклетки. Оставив тело гидры, спермато-зоиды по воде следуют к другим особям. Отыскав яйцеклетки, они оплодотворяют их. Образуется зигота, вокруг которой возникает плот-ная оболочка. Это оплодотворенное яйцо остается в теле гидры. Обычно половое размножение происходит осенью. Зимой взрослые гидры по-гибают, а яйца переживают зиму на дне водоема. Весной зигота начина-ет делиться, образуя два слоя клеток. Из них и развивается маленькая гидра.
На этой странице материал по темам:
Процессы жизнедеятельности гидры
Жизнедеятельность гидры сокращёное сочинение
Назовите сходные черты и отличия в строении гидры и губки
Размножение гидры. регенерация. значение в природе.
Почему название подошва одинаково для губки для гидры
Вопросы по этому материалу: